本网讯(通讯员 孙航军)在经典植物免疫学中,分子模式触发的免疫反应(PTI)和效应子触发的免疫反应(ETI)共同构成了植物的先天免疫系统。两者协同参与植物对病原真菌与细菌的识别与防御。然而,由于病毒的胞内专性寄生特性及其非细胞特性,PTI是否参与植物的抗病毒防御一直存在争议。
近日,澳门新葡京娱乐城 李洪连教授团队|孙航军副教授、孙炳剑教授在国际著名期刊Plant Cell & Environment上发表了题为“Unraveling Plant-Virus Warfare: Pattern-Triggered Immunity-Mediated Antiviral Defense and Viral Evasion Strategies”的综述论文。该论文详细介绍了PTI核心组分(膜受体蛋白PRR、MAPK激酶、抗病激素)的抗病毒作用,以及病毒对这些组分的干扰机制。同时,论文探讨了植物可能具备的识别病毒的机制,以及PTI末端输出物质对病毒侵染的抑制作用。
在经典的PTI信号途径中,植物膜受体蛋白的胞外结构域识别病原细菌和真菌的保守分子模式(PAMP),随后构象发生变化并与共受体形成复合体,通过相互或自我磷酸化激活胞内激酶结构域。胞内激酶结构域直接或间接激活下游的RLCK激酶、MAPK激酶,进而诱导抗病激素水杨酸、茉莉酸、乙烯的合成。经过复杂的信号转导和转录重排,最终诱导植物产生大量抗病物质。这些物质中,有些能直接杀死病原细胞,例如植保素、病程相关蛋白;另一些则通过增强细胞壁的机械强度,阻止病原真菌或细菌的侵入与扩散,例如木质素、胼胝质等。这些信号转导途径的激活和抗病物质的产生是PTI的保守下游反应,对几乎所有病原细菌和真菌均具有抑制作用。
然而,与其它病原物不同,病毒是一种非细胞生物,在寄主细胞外不进行代谢反应,仅通过病毒核酸和衣壳蛋白维持稳定性,因此被视为非生命实体。理论上,植保素等直接杀伤病原细胞的抗病物质无法有效“杀死”病毒颗粒。此外,植物病毒不需主动侵入细胞,而是借助昆虫取食造成的伤口或机械损伤造成的微伤口被动进入植物细胞。并且一旦进入植物细胞,病毒可以通过胞间连丝扩散至整个植株,无需离开细胞。因此PTI诱导的木质素沉积等细胞壁抗性似乎对病毒无效。这些问题引发了人们对PTI免疫途径是否具有抗病毒作用的质疑。
近日,澳门新葡京娱乐城 李洪连教授团队|孙航军副教授、孙炳剑教授在国际著名期刊Plant Cell & Environment上发表了题为“Unraveling Plant-Virus Warfare: Pattern-Triggered Immunity-Mediated Antiviral Defense and Viral Evasion Strategies”的综述论文。该论文详细介绍了PTI核心组分(膜受体蛋白PRR、MAPK激酶、抗病激素)的抗病毒作用,以及病毒对这些组分的干扰机制。同时,论文探讨了植物可能具备的识别病毒的机制,以及PTI末端输出物质对病毒侵染的抑制作用。
在经典的PTI信号途径中,植物膜受体蛋白的胞外结构域识别病原细菌和真菌的保守分子模式(PAMP),随后构象发生变化并与共受体形成复合体,通过相互或自我磷酸化激活胞内激酶结构域。胞内激酶结构域直接或间接激活下游的RLCK激酶、MAPK激酶,进而诱导抗病激素水杨酸、茉莉酸、乙烯的合成。经过复杂的信号转导和转录重排,最终诱导植物产生大量抗病物质。这些物质中,有些能直接杀死病原细胞,例如植保素、病程相关蛋白;另一些则通过增强细胞壁的机械强度,阻止病原真菌或细菌的侵入与扩散,例如木质素、胼胝质等。这些信号转导途径的激活和抗病物质的产生是PTI的保守下游反应,对几乎所有病原细菌和真菌均具有抑制作用。
然而,与其它病原物不同,病毒是一种非细胞生物,在寄主细胞外不进行代谢反应,仅通过病毒核酸和衣壳蛋白维持稳定性,因此被视为非生命实体。理论上,植保素等直接杀伤病原细胞的抗病物质无法有效“杀死”病毒颗粒。此外,植物病毒不需主动侵入细胞,而是借助昆虫取食造成的伤口或机械损伤造成的微伤口被动进入植物细胞。并且一旦进入植物细胞,病毒可以通过胞间连丝扩散至整个植株,无需离开细胞。因此PTI诱导的木质素沉积等细胞壁抗性似乎对病毒无效。这些问题引发了人们对PTI免疫途径是否具有抗病毒作用的质疑。
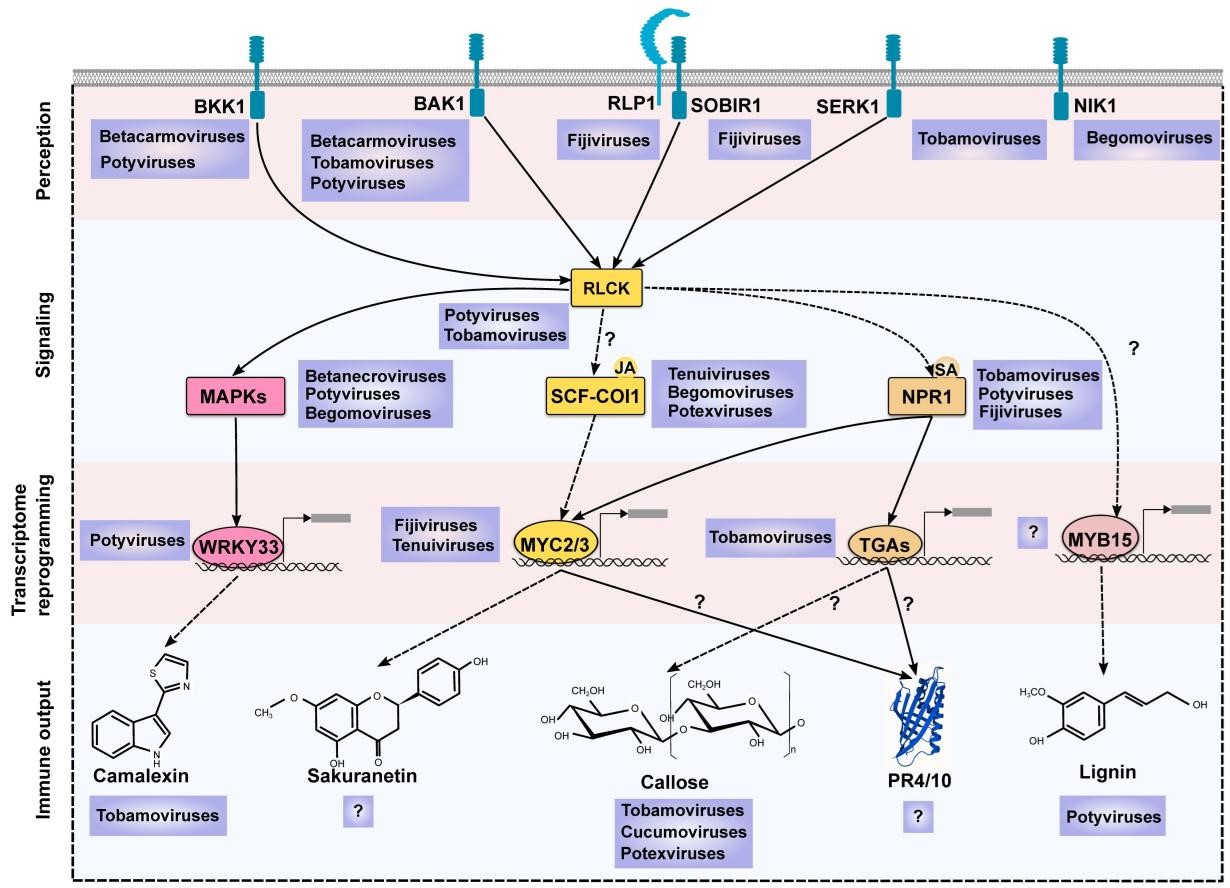
图1. PTI核心组分的抗病毒作用
作者系统总结了近年来对PTI核心组分抗病毒作用的研究进展。大量研究报道了多种PTI下游信号通路(如MAPK途径、水杨酸途径、茉莉酸途径)可被不同病毒激活。利用基因敲除或过表达转基因遗传材料,人们也证实这些保守途径确实参与植物的抗病毒防御体系(图1)。此外,与病原真菌和细菌相似,病毒在与植物长期的“军备竞赛”过程中也进化出了自身的效应蛋白,能够通过靶向PTI核心组分干扰PTI信号转导。例如,膜受体蛋白、MAPK激酶(图2)、水杨酸/茉莉酸合成途径、SCF-COI1复合体、MYCs转录因子、NPR1、TGAs转录因子等(图3),均为病毒的重点干扰靶标。作者认为这些研究结果足够支撑PTI是植物抗病毒免疫系统关键组成部分的观点。

图2 病毒干扰膜受体蛋白和MAPK途径的策略

图3 病毒干扰水杨酸和茉莉酸途径的策略
尽管如此,仍有两个核心问题尚未得到解答。首先,病毒的保守分子模式(PAMP)有哪些?植物又是如何感知病毒的分子模式并激发PTI免疫系统的?作者认为,病毒的胞外粒体结构并不具备保守性。相反,病毒的保守性往往体现在胞内复制时产生的结构和物质,例如RNA病毒复制时都会产生囊泡状结构以及双链RNA分子。这些物质和结构可能是病毒的PAMP。由于病毒一旦进入植物细胞就可通过胞间连丝扩散至整个植株,无需再次离开和侵入细胞,因此植物对病毒的识别过程可能存在于胞内而不是细胞膜上(图4)。然而,只有鉴定到能够直接识别病毒PAMP的受体蛋白,并解析受体蛋白如何通过构象变化激活下游途径,才能真正回答这个问题。
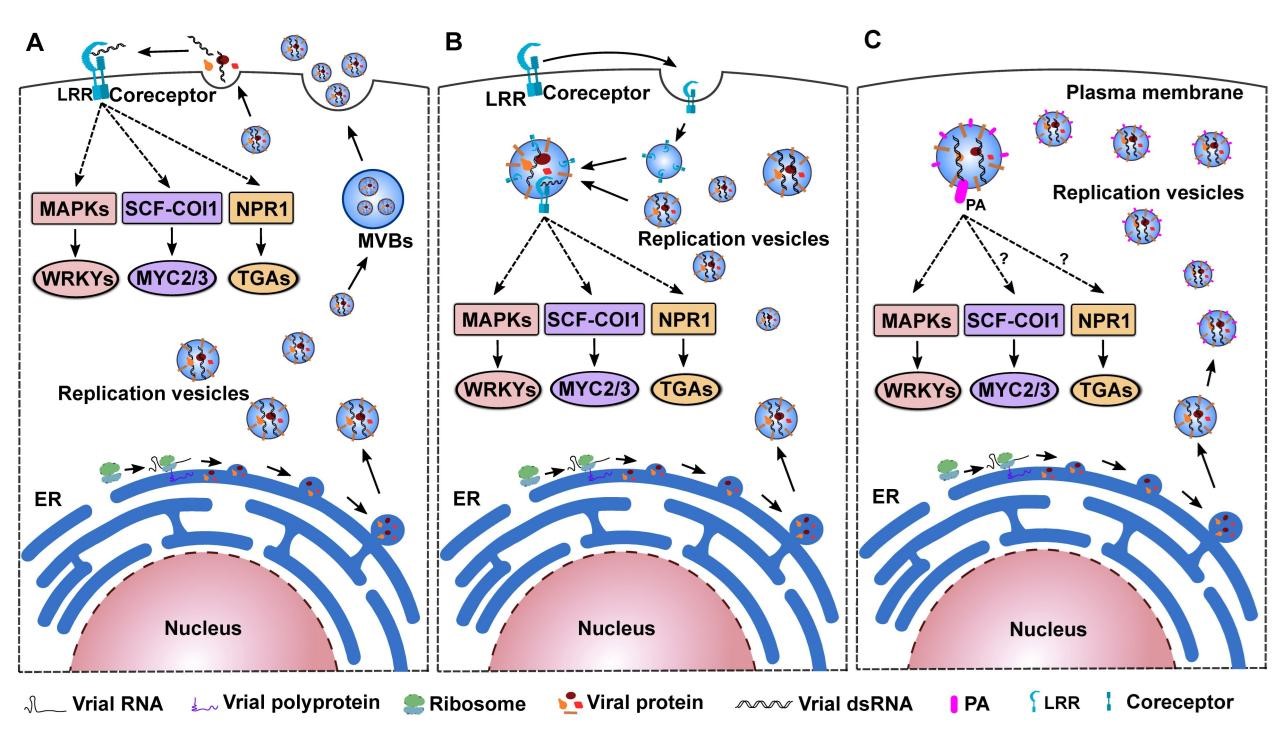
图4 植物识别病毒的潜在机制
其次,在PTI诱导产生的抗病物质中,哪些组分真正起着抵抗病毒侵染的作用?目前较为认可的是胼胝质能通过堵塞胞间连丝限制病毒的系统侵染。文中讨论了其它物质的抗病毒机制。例如,某些植保素(如Camalexin)可能具有干扰病毒外壳蛋白组装的功能,而一些病程相关蛋白(如PR4、PR10)可能具有降解病毒核酸的功能,木质素可能通过增强导管的木质化程度限制病毒的系统侵染。此外,PTI还可能产生一些尚未被鉴定到的抗病毒物质。但这些猜测仍需要更多研究验证。
河南农业大学孙航军副教授和孙炳剑教授为论文的共同通讯作者,李洪连教授对本论文提供了重要的指导。本研究得到国家自然科学基金(31901855)、河南省高校科技创新团队(25IRTSTHN031)、海南省烟草公司科技项目(2022002)的资助。
论文链接://onlinelibrary.wiley.com/doi/10.1111/pce.70028
河南农业大学孙航军副教授和孙炳剑教授为论文的共同通讯作者,李洪连教授对本论文提供了重要的指导。本研究得到国家自然科学基金(31901855)、河南省高校科技创新团队(25IRTSTHN031)、海南省烟草公司科技项目(2022002)的资助。
论文链接://onlinelibrary.wiley.com/doi/10.1111/pce.70028





